Salientia
Frogs and toads
David Cannatella



This tree diagram shows the relationships between several groups of organisms.
The root of the current tree connects the organisms featured in this tree to their containing group and the rest of the Tree of Life. The basal branching point in the tree represents the ancestor of the other groups in the tree. This ancestor diversified over time into several descendent subgroups, which are represented as internal nodes and terminal taxa to the right.

You can click on the root to travel down the Tree of Life all the way to the root of all Life, and you can click on the names of descendent subgroups to travel up the Tree of Life all the way to individual species.
For more information on ToL tree formatting, please see Interpreting the Tree or Classification. To learn more about phylogenetic trees, please visit our Phylogenetic Biology pages.
close boxIntroduction
Salientia includes frogs and toads, and their close fossil relatives. The closest living relatives of the Salientia are the other amphibians, salamanders and caecilians. Frogs outnumber both of these groups substantially; as of May 2007, there are 172 living species of caecilians, 557 living salamanders, and 5424 living species of frogs.
The earliest known salientian is †Triadobatrachus massinoti, from the Early Triassic of Madagascar. This "proto-frog" is about 250 million years old. "Proto-frog" refers to the fact that it had not yet quite evolved the combination of features that are typically associated with frogs. For more information see †Triadobatrachus massinoti.
The earliest "true" frogs include †Prosalirus bitis and †Vieraella herbsti, from the Early Jurassic era. Thus, perfectly respectable frogs were around just before most of the major groups of dinosaurs had appeared. †Notobatrachus degiustoi from the Middle Jurassic is just a bit younger, about 155-170 million years old.
There are many distinctive features of living frogs. Frogs have at most nine vertebrae in front of the sacrum, and the three or four posterior to the sacrum are fused into a rod called the urostyle. In contrast, caecilians and salamanders have many more vertebrae and they do not have a urostyle. Frogs do not possess tails in the adult stage ("Anura" means without tail), as caecilians and salamanders do. Frogs also have a radioulna, which represents a fused radius and ulna (bones of the forearm), and a tibiofibula, the fused tibia and fibula (bones of the shank). The tibiale and fibulare (ankle bones; also called astragalus and calcaneum) of frogs are greatly elongate. Thus there is effectively an additional lever system that frogs can utilize in jumping. Indeed, the origin of saltation and its morphological correlates (lack of a tail, reduction in vertebrae, elongation of propulsive segments of the body) seems to be one of the features that clearly sets frogs apart from other major vertebrate groups (Gans and Parsons, 1966).
In addition to morphological distinctions, frogs also have a distinctive life phase known as the tadpole, which is a highly specialized "eating machine." Salamanders and caecilians have a larval form, but in neither does the larva possess the many specializations (such as the ceratohyal pump) that frog tadpoles have (Wassersug, 1974). Even the most basal living frogs have the beginnings of a unique mechanism of tongue projection (Nishikawa and Cannatella, 1991; Nishikawa and Roth, 1991) that is associated with extreme modification of the gill arches into a fused hyobranchial plate.
Although there is no scientific distinction between "frogs" and "toads", frogs are typically smooth-skinned, have long hind limbs for leaping, and live in water, while toads have warty, drier skin, with shorter hind limbs for hopping, and live on land (Halliday and Adler, 1986).
With so many species of frogs and toads, it is not surprising that they inhabit a wide variety of habitats. Habitat types range from arid desert regions to mountainous regions to swamps to tropical rainforests.
Temperature and water regulation are critical to frogs and toads, and amphibians in general. Being ectotherms, frogs and toads are reliant on the ambient temperature for body temperature regulation. In the winter months, frogs in temperate zones cannot remain active and must enter into a state of torpor, or extremely reduced activity. In the contrasting summer months, frogs can avoid the extreme heat by remaining underground in daylight, and being active at night (Halliday and Adler, 1986).
Salientians are also susceptible to the loss of body water due to extremely hot or dry conditions. Moisture regulation in frogs varies with their habitat. Those in temperate climates maintain moist skin to aid in evaporative cooling. As external air passes over the moist skin, the frog's body temperature is lowered. Additionally, permeable skin allows the frog the ability to absorb water simply by jumping into a pond or sitting in a puddle. Frogs in arid regions, on the other hand, have different ways of regulating body water. Their skin is often impermeable to water to prevent rapid evaporation and dehydration. Instead, they may cover their bodies with a thick mucus, or burrow to avoid the heat altogether.
Breeding in frogs is triggered by environmental cues such as temperature change and rainfall. During the breeding season (which varies with each species) hundreds or thousands of frogs may be seen in a congregation. Male frogs attract mates by calling; often many males call in chorus. Calling usually occurs near a body of water, such as a pond, where the eggs can be laid and fertilized. Egg masses may be laid in long chains or in large clumps. Parental care in frogs and toads is variable; some species lay many smaller eggs and have little parental care, while others lay a few larger eggs and remain with them until tadpoles or froglets develop.
Geographic Distribution
The global distribution of Salientia is indicated in red.
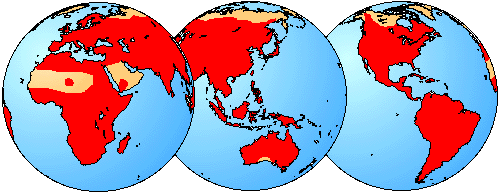
Herpetological Fauna of Texas
The Salientia clade includes all frogs and toads from around the world. For a list of only the frog species found in Texas, as well as other amphibians and reptiles, see Herps of Texas
Discussion of Phylogenetic Relationships
The name Salientia generally has referred to †Triadobatrachus + Anura (Milner, 1988). Ford and Cannatella (1993) defined it as a stem-based name for amphibians that are more closely related to Anura than to Caudata or Gymnophiona. Synapomorphies that unite all of the currently known taxa in Salientia include 14 presacral vertebrae, elongate and anteriorly directed ilium, presence of a frontoparietal, and a toothless dentary (Milner, 1988). To these, Trueb and Cloutier (1991) added the absence of a lacrimal and unicapitate ribs as other unique synapomorphies, and four other synapomorphies that showed homoplasy among closely related dissorophoid temnospondyls.
References
Click here for more detailed references.
Báez, A. M., and N. G. Basso. 1996. The earliest known frogs of the Jurassic of South America: review and cladistic appraisal of their relationships. Münchner Geowiss. Abh. (a) 30:131-158.
Cannatella, D. C. 1985. A phylogeny of primitive frogs (archaeobatrachians). Ph.D. Dissertation, The University of Kansas, Lawrence.
Cannatella, D. C. 1989. On the monophyly of discoglossoid frogs. Pp. 230-231 In H. Splechtna and H. Hilgers (Eds.), Trends in Vertebrate Morphology. Gustav Fischer Verlag, Stuttgart.
Cannatella, D. C., and D. M. Hillis. 1993. Amphibian phylogeny: phylogenetic analysis of morphology and molecules. Herpetol. Monogr. 7:1-7.
de Queiroz, K., and J. Gauthier. 1992. Phylogenetic taxonomy. Ann. Rev. Ecol. Syst. 23:449-80.
Dubois, A. 1983. Classification et nomenclature supragénérique des amphibiens anoures. Bull. Soc. Linn. Lyon 52:270-276.
Dubois, A. 1984. La nomenclature supragénérique des amphibiens anoures. Mémoires Museum Nat. d'Histoire Naturelle, Ser.A 131:1-64.
Duellman, W. E. 1975. On the classification of frogs. Occ. Pap. Mus. Nat. Hist. Univ. Kansas (42):1-15.
Duellman, W. E., and L. Trueb. 1986. Biology of Amphibians. McGraw-Hill Book Co., New York.
Estes, R., and O. A. Reig. 1973. The early fossil record of frogs: a review of the evidence. Pp. 11-63 In J. L. Vial (Ed.), Evolutionary Biology of the Anurans: Contemporary Research on Major Problems. University of Missouri Press, Columbia.
Ford, L. S. 1989. The phylogenetic position of poison-dart frogs (Dendrobatidae): reassessment of the neobatrachian phylogeny with commentary on complex character systems. Ph.D. Dissertation, The University of Kansas, Lawrence, Kansas.
Ford, L. S., and D. C. Cannatella. 1993. The major clades of frogs. Herp. Monogr. 7:94-117.
Frost, D. R. (Ed.) 1985. Amphibian Species of the World. Allen Press and the Association of Systematics Collections, Lawrence, Kansas.
Gans, C., and T. S. Parsons. 1966. On the origin of the jumping mechanism in frogs. Evolution 20(1):92-99.
Griffiths, I. 1963. The phylogeny of the Salientia. Biological Reviews 38:241-292.
Halliday, T. R., and K. Adler (Eds.). 1987. The Encyclopedia of Reptiles and Amphibians. Equinox, Oxford.
Hay, J. M., I. Ruvinsky, S. B. Hedges, and L. R. Maxson. 1995. Phylogenetic relationships of amphibian families inferred from DNA sequences of mitochondrial 12S and 16S ribosomal RNA genes. Mol. Biol. Evol. 12(5):928-937.
Hedges, S. B., K. D. Moberg, and L. R. Maxson. 1990. Tetrapod phylogeny inferred from 18s and 28s ribosomal RNA sequences and a review of the evidence for amniote relationships. Mol. Biol. Evol. 7:607-633.
Hedges, S. B. and L. R. Maxson. 1993. A molecular perspective on lissamphibian phylogeny. Herpetol. Monogr. 7:27-42.
Hillis, D. M., L. K. Ammerman, M. T. Dixon, and R. O. de Sá. 1993. Ribosomal DNA and the phylogeny of frogs. Herpetol. Monogr. 7:118-131.
Inger, R. F. 1967. The development of a phylogeny of frogs. Evolution 21:369-384.
Kluge, A. G., and J. S. Farris. 1969. Quantitative phyletics and the evolution of anurans. Syst. Zool. 18:1-32.
Lanza, B., J. M. Cei, and E. G. Crespo. 1976. Further immunological evidence for the validity of the family Bombinidae (Amphibia, Salientia). Monit. Zool. Ital. (NS) 10:311-314.
Laurent, R. F. 1979. Esquisse d'une phylogenèse des anoures. Bull. Soc. Zool. France 104:397-422.
Laurent, R. F. 1986. Sous classe des lissamphibiens. Systématique. Pp. 594-797 In P.-P. Grassé and M. Delsol (Eds.), Traité de Zoologie, Tome 14, 1B. Masson, Paris.
Lynch, J. D. 1971. Evolutionary relationships, osteology, and zoogeography of leptodactyloid frogs. Misc. Publ. Mus. Nat. Hist. Univ. Kansas (53):531-238.
Lynch, J. D. 1973. The transition from archaic to advanced frogs. Pp. In J. L. Vial (Ed.), Evolutionary Biology of the Anurans: Contemporary Research on Major Problems. University of Missouri Press, Columbia.
Milner, A. R. 1988. The relationships and origin of living amphibians. Pp. 59-102 In M. J. Benton (Ed.), The Phylogeny and Classification of the Tetrapods. 1. Amphibians, Reptiles, Birds. Oxford University Press, Oxford.
Milner, A. R. 1993. The Paleozoic relatives of lissamphibians. Herpetol. Monogr. 7:8-27.
Noble, G. K. 1922. The phylogeny of the Salientia. I. The osteology and the thigh musculature; their bearing on classfication and phylogeny. Bull. Amer. Mus. Nat. Hist. 46:1-87.
Noble, G. K. 1931. The Biology of the Amphibia. McGraw-Hill, New York.
Orton, G. 1957. The bearing of larval evolution on some problems in frog classification. Syst. Zool. 6:79-86.
Peters, J. A. 1964. Dictionary of Herpetology. Hafner Publishing Co., New York.
Pusey, H. K. 1943. On the head of the liopelmid frog, Ascaphus truei. I. The chondrocranium, jaws, arches, and muscles of a partly grown larva. Quart. J. Micr. Sci. 84:106-185.
Reig, O. A. 1958. Proposiciones para una nueva macrosistematica de los anuros. Nota preliminar. Physis 21:109-118.
Rocek, Z. 1981. Cranial anatomy of frogs of the family Pelobatidae Stannius, 1856, with outlines of their phylogeny and systematics. Acta Univ. Carolinae Biol. 1980:1-164.
Saint-Aubain, M. L. de 1981. Amphibian limb ontogeny and its bearing on the phylogeny of the group. Zeit. Zool. Systematik u. Evolutionsforschung 19:175-194.
Savage, J. M. 1973. The geographic distribution of frogs: patterns and predictions. Pp. 351-445 In J. L. Vial (Ed.), Evolutionary Biology of the Anurans: Contemporary Research on Major Problems. University of Missouri Press, Columbia.
Slabbert, G. K., and W. A. Maree. 1945. The cranial morphology of the Discoglossidae and its bearing upon the phylogeny of the primitive Anura. Ann. Univ. Stellen. 23a:91-97.
Sokol, O. M. 1975. The phylogeny of anuran larvae: a new look. Copeia 1975:1-24.
Sokol, O. M. 1977a. A subordinal classification of frogs (Amphibia: Anura). J. Zool., London 182:505-508.
Starrett, P. H. 1968. The phylogenetic significance of the jaw musculature in anuran amphibians. PhD. Dissertation. University of Michigan, Ann Arbor, Michigan.
Starrett, P. H. 1973. Evolutionary patterns in larval morphology. Pp. 251-271. In J. L. Vial (Ed.), Evolutionary Biology of the Anurans: Contemporary Research on Major Problems. University of Missouri Press, Columbia.
Stephenson, E. M. T. 1951. The anatomy of the head of the New Zealand frog, Leiopelma. Trans. Zool. Soc. London 27:255-305.
Trueb, L. 1973. Bones, frogs, and evolution. Pp. In (Eds.), In: Evolutionary Biology of the Anurans: Contemporary Research on Major Problems (J.L. Vlal, ed.), pp. 65-132. Univ. of Missouri Press, Columbia.
Trueb, L., and R. Cloutier. 1991. A phylogenetic investigation of the inter- and intrarelationships of the Lissamphibia (Amphibia: Temnospondyli). Pp. 233-313. In H.-P. Schultze and L. Trueb (Eds.), Origins of the Higher Groups of Tetrapods. Cornell University Press, Ithaca.
Trueb, L. 1993. Patterns of cranial diversity among the Lissamphibia. Pp. 255-343 In J. Hanken and B. Hall (Eds.) The Vertebrate Skull. University of Chicago Press, Chicago.
Information on the Internet
- AmphibiaTree. The mission of this project is the realization of a comprehensive phylogeny of all taxa of extant and extinct amphibians.
- AmphibiaWeb. A site inspired by global amphibian declines, is an online system that allows free access to information on amphibian biology and conservation.
- Amphibian Species of the World. American Museum of Natural History.
- FrogWeb: Amphibian Declines & Deformities. National Biological Information Infrastructure.
- IUCN Amphibian Specialist Group
- FROGS.ORG. Website of the Amphibian Conservation Alliance.
- Frogs. A Chorus of Colors. American Museum of Natural History.
- Exploratorium: Frogs. An exhibition at the Exploratorium, the museum of science, art and human perception.
- The Frog Files. Information on the frogs of Western Australia.
- MidWest Frogs. Frog call video clips.
- Frogs Home. University of Wisconsin Sea Grant Institute.
- A Thousand Friends of Frogs. Connecting children, parents, educators, and scientists to study and celebrate frogs and their habitats. Center for Global Environmental Education, Hamline University.
- Frogs and Toads in Color and Sound. Lang Elliott, NatureSound Studio.
- Manitoba Frog and Toad Calls.
- Virtual frog dissections:
- Froguts. The first true virtual online frog dissection.
- LBL ITG Whole Fro Project. Interactive frog dissection kit.
Title Illustrations

| Scientific Name | Ascaphus truei |
|---|---|
| Location | Oregon |
| Specimen Condition | Live Specimen |
| Sex | Male |
| Image Use |
 This media file is licensed under the Creative Commons Attribution-ShareAlike License - Version 3.0. This media file is licensed under the Creative Commons Attribution-ShareAlike License - Version 3.0.
|
| Copyright |
© 1995 David Cannatella

|
About This Page
Original production of the Salientia clade of the Tree of Life was funded in part by Project QUEST, the Texas Memorial Museum, and the College of Natural Sciences of the University of Texas, Austin, Texas. Updates funded by the National Science Foundation, through the Assembling the Tree of Life program.
David Cannatella
University of Texas, Austin, Texas, USA
Correspondence regarding this page should be directed to David Cannatella at
Page copyright © 1997 David Cannatella
 Page: Tree of Life
Salientia. Frogs and toads.
Authored by
David Cannatella.
The TEXT of this page is licensed under the
Creative Commons Attribution License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
Page: Tree of Life
Salientia. Frogs and toads.
Authored by
David Cannatella.
The TEXT of this page is licensed under the
Creative Commons Attribution License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
- Content changed 11 January 2008
Citing this page:
Cannatella, David. 2008. Salientia. Frogs and toads. Version 11 January 2008 (under construction). http://tolweb.org/Salientia/14938/2008.01.11 in The Tree of Life Web Project, http://tolweb.org/